Masha Gessen first came to my attention because of the controversy caused by her being awarded the Hannah Arendt Prize in 2023. A week before the planned award ceremony Gessen published an essay in the New Yorker with the title ‘In the Shadow of the Holocaust’. In this essay she portrays the Israeli military actions in the Gaza Strip as being comparable to Nazi liquidations of Jewish ghettos. I have written about this issue elsewhere (in German) and I will say no more about it here. While trying to find out more about this person I discovered that she had written a biography of Grigori Perelman with the title ‘Perfect Rigor’. Perelman is one of the most remarkable mathematicians of recent years. He resolved one of the most famous open problems in mathematics, the Poincaré conjecture, formulated in 1904. For this he was offered a Fields medal in 2006, the most prestigious award in mathematics, which he refused. Later, in 2010, he was offered a one million dollar prize by the Clay Foundation, which he also refused, despite living in poverty. As I mentioned in previous posts here and here I had the privilege to hear the first lecture which Perelman gave on his proof of the Poincare conjecture and afterwards I went to a cafe with him together with colleagues, so that I was able to get a further personal impression of him. Masha Gessen was never able to talk to Perelman because by the time she was working on her book he had completely withdrawn from public contacts. She was, however, able to talk to people who had known Perelman very well. She also has special insights into the world in which Perelman grew up in the Soviet Union since she grew up in the same society.
Masha Gessen is not a mathematician although she did apparently have a good school education in mathematics. For that reason her ability to write about the Poincaré conjecture itself is limited. What she does write on that is imprecise in a way which is likely to be a little irritating for people who know the subject fairly well and could be confusing for professional mathematicians with no special knowledge of the subject. However that is only a small part of the book. The main subjects are Perelman himself, the people around him and the society around him. In the previous post where I mentioned Perelman I compared him to another famous mathematician who refused prestigious prizes, Grothendieck. Now I still see similarities between the two men but also major differences. Grothendieck was a very political person and his motivation for refusing prizes probably came mainly from politics. Perelman, as he is portrayed in the book, is very different. He sees it as a disgrace for a mathematician to pay attention to politics. For him mathematics is much more important and valuable than politics. His reasons for refusing prizes are rather personal. He is absolutely convinced of the value of his work and he is not prepared to accept that others (who understand less about the subject than himself) should judge it. The book was written before Perelman had been awarded the Clay prize but at that time it already seemed probable that he would not accept it. At that time his complaint was that it should have been given equally to him and Richard Hamilton, on whose work he had built. However other examples are given where Perelman refuses something for a certain reason, the offer is modified to remove this obstacle and he still refuses. It seems that in a way Perelman was an exceptionally honest person but that he often kept his motives hidden. I notice that I am talking about Perelman here in the past tense and the reason is that he long ago cut off all contact with the rest of the world, with the exception of his mother. It should be mentioned that his mother herself wanted to become a professional mathematician but was prevented from doing so by antisemitism and sexism.
An interesting aspect of the book for me is the picture it gives of mathematics in the Soviet Union, something which was previously just a black box for me. Another is that I know some of the people who play a role in the book personally and I also experienced some aspects of the ICM in Madrid described in the book myself, for instance the lecture of Hamilton, and it is interesting to have some of the background to what I saw revealed. I did not specially like the way the book is written but it certainly kept my attention and I did learn a lot.
 from
from ![[-1,1]](https://s0.wp.com/latex.php?latex=%5B-1%2C1%5D&bg=ffffff&fg=333333&s=0&c=20201002) to the real line which is non-zero at both endpoints. If its values at both endpoints have the same sign the degree is zero. If
to the real line which is non-zero at both endpoints. If its values at both endpoints have the same sign the degree is zero. If  and
and  then the degree is one. If
then the degree is one. If  and
and  then it is minus one. In this situation it is clear that the degree is unchanged by continuous deformations of
then it is minus one. In this situation it is clear that the degree is unchanged by continuous deformations of 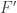 at the zeroes of
at the zeroes of 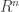 to itself has a fixed point. Another replaces the ball by an arbitrary non-empty compact convex set. I will not discuss here how the second, apparently stronger, version follows from the first. It is the second version which is extended in the Schauder fixed point theorem. That theorem says that a compact mapping of a non-empty closed, bounded, convex subset of a Banach space to itself has a fixed point. Here a compact mapping is a continuous one which maps bounded sets to compact sets. In an alternative version of the theorem it is only assumed that the mapping is continuous but it is assumed that the set is compact. This fixed point theorem can be used to develop Leray-Schauder theory. One version of this, discussed in Zeidler’s book, is as follows. Let
to itself has a fixed point. Another replaces the ball by an arbitrary non-empty compact convex set. I will not discuss here how the second, apparently stronger, version follows from the first. It is the second version which is extended in the Schauder fixed point theorem. That theorem says that a compact mapping of a non-empty closed, bounded, convex subset of a Banach space to itself has a fixed point. Here a compact mapping is a continuous one which maps bounded sets to compact sets. In an alternative version of the theorem it is only assumed that the mapping is continuous but it is assumed that the set is compact. This fixed point theorem can be used to develop Leray-Schauder theory. One version of this, discussed in Zeidler’s book, is as follows. Let 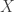 be a Banach space and
be a Banach space and  a compact mapping. If all solutions of the equation
a compact mapping. If all solutions of the equation  with
with  satisfy
satisfy  for a constant
for a constant  then
then 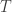 has a fixed point. Another nice discussion of these topics can be found in the standard textbook on elliptic equations by Gilbarg and Trudinger. They also discuss a more general version of the Leray-Schauder theory where the mapping
has a fixed point. Another nice discussion of these topics can be found in the standard textbook on elliptic equations by Gilbarg and Trudinger. They also discuss a more general version of the Leray-Schauder theory where the mapping  is replaced by a more general one. In other words the requirement of the linear dependence of the mapping on the parameter is dropped. Here we see the relation to degree theory. They present quite self-contained proofs of these results, modulo the Brouwer fixed point theorem. In fact they do not need that theorem for a general compact convex subset, but only the case of the convex hull of a finite set. The theory can be used to obtain existence theorems for boundary value problems for nonlinear elliptic equations (or initial boundary value problems for nonlinear parabolic equations). The necessary input is a priori estimates together with some compactness. The latter comes from the regularizing properties of solving equations of these kinds. The idea then is to deform the problem to be solved continuously to one which is easier to solve, or even trivial.
is replaced by a more general one. In other words the requirement of the linear dependence of the mapping on the parameter is dropped. Here we see the relation to degree theory. They present quite self-contained proofs of these results, modulo the Brouwer fixed point theorem. In fact they do not need that theorem for a general compact convex subset, but only the case of the convex hull of a finite set. The theory can be used to obtain existence theorems for boundary value problems for nonlinear elliptic equations (or initial boundary value problems for nonlinear parabolic equations). The necessary input is a priori estimates together with some compactness. The latter comes from the regularizing properties of solving equations of these kinds. The idea then is to deform the problem to be solved continuously to one which is easier to solve, or even trivial.