In my previous post on this subject I discussed the question of the status of the variables in the Poolman model of photosynthesis and in the end I was convinced that I had understood which concentrations are to be considered as dynamical unknowns and which as constants. The Poolman model is a modified version of the Pettersson model and the corresponding questions about the nature of the variables have the same answers in both cases. What I am calling the Pettersson model was introduced in a paper of Pettersson and Ryde-Pettersson (Eur. J. Biochem 175, 661) and there the description of the variables and the equations is rather complete and comprehensible. Now I will go on to consider the second question raised in the previous post, namely what the evolution equations are. The evolution equations in the Poolman model are modifications of those in the Pettersson model and are described relative to those in the original paper on the former model. For this reason I will start by describing the equations for the Pettersson model. As a preparation for that I will treat a side issue. In a reaction network a reaction whose rate depends only on the concentrations of the substances consumed in the reaction is sometimes called NAC (for non-autocatalytic). For instance this terminology is used in the paper of Kaltenbach quoted in the previous post. The opposite of NAC is the case where the reaction rate is modulated by the concentrations of other substances, such as activators or inhibitors.
The unknowns in the Pettersson model are concentrations of substances in the stroma of the chloroplast. The substances involved are 15 carbohydrates bearing one or more phosphate groups, inorganic phosphate and ATP, thus 17 variables in total. In addition to ordinary reactions between these substances there are transport processes in which sugar phosphates are moved from the stroma to the cytosol in exchange for inorganic phosphate. For brevity I will also refer to these as reactions. The total amount of phosphate in the stroma is conserved and this leads to a conservation law for the system of equations, a fact explicitly mentioned in the paper. On the basis of experimental data some of the reactions are classified as fast and it is assumed that they are already at equilibrium. They are also assumed to be NAC and to have mass-action kinetics. This defines a set of algebraic equations. These are to be used to reduce the 17 evolution equations which are in principle there to five equations for certain linear combinations of the variables. The details of how this is done are described in the paper. I will now summarize how this works. The time derivatives of the 16 variables other than inorganic phosphate are given in terms of linear combinations of 17 reaction rates. Nine of these reaction rates, which are not NAC, are given explicitly. The others have to be treated using the 11 algebraic equations coming from the fast reactions. The right hand sides 
Consider now the Poolman model. One way in which it differs from the Pettersson model is that starch degradation is included. The other is that while the kinetics for the ‘slow reactions’ (i.e. those which are not classified as fast in the Pettersson model) are left unchanged, the equilibrium assumption for the fast reactions is dropped. Instead the fast reactions are treated as reversible with mass action kinetics. In the thesis of Sergio Grimbs (Towards structure and dynamics of metabolic networks, Potsdam 2009) there is some discussion of the models of Poolman and Pettersson. It is investigated whether information about multistability in these models can be obtained using ideas coming from chemical reaction network theory. Since the results from CRNT considered require mass action kinetics it is implicit in the thesis that the systems are being considered which are obtained by applying mass action to all reactions in the networks for the Poolman and Pettersson models. These systems are therefore strictly speaking different from those of Pettersson and Poolman. In any case it turned out that these tools were not useful in this example since the simplest results did not apply and for the more complicated computer-assisted ones the systems were too large.
In the Pettersson paper the results of computations of steady states are presented and a comparison with published experimental results looks good in a graph presented there. So whay can we not conclude that the problem of modelling the dynamics of the Calvin cycle was pretty well solved in 1988? The paper contains no details on how the simulations were done and so it is problematic to repeat them. Jablonsky et. al. set up simulations of this model on their own and found results very different from those reported in the original paper. In this context the advantage of the Poolman model is that it has been put into the BioModels database so that the basic data is available to anyone with the necessary experience in doing simulations for biochemical models. Forgetting the issue of the reliability of their simulations, what did Petterson and Ryde-Pettersson find? They saw that depending on the external concentration of inorganic phosphate there is either no positive stationary solution (for high values of this parameter) or two (for low values) with a bifurcation in between. When there are two stationary solutions one is stable and one unstable. It looks like there is a fold bifurcation. There is a trivial stationary solution with all sugar concentrations zero for all values of the parameter. When the external phosphate concentration tends to zero the two positive stationary solutions coalesce with the trivial one. The absence of positive stationary solutions for high phosphate concentrations is suggested to be related to the concept of ‘overload breakdown’. This means that sugars are being exported so fast that the production from the Calvin cycle cannot keep up and the whole system breaks down. It would be nice to have an analytical proof of the existence of a fold bifurcation for the Pettersson model but that is probably very difficult to get.
 to vertex
to vertex  if there is any set of concentrations for which the corresponding partial derivative is non-zero and instead of being labelled with a sign the edge is labelled with a function, namely the partial derivative itself.
if there is any set of concentrations for which the corresponding partial derivative is non-zero and instead of being labelled with a sign the edge is labelled with a function, namely the partial derivative itself.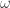 -limit points where some concentration vanishes. This property is sometimes known as persistence.
-limit points where some concentration vanishes. This property is sometimes known as persistence.