There is evidence to suggest that oscillations in levels of calcium inside and outside cells are used as a signalling mechanism. A variety of mathematical models have been introduced to study this phenomenon. Here I will discuss some aspects of the subject. A more general review can be found in this Scholarpedia article. In the plasma membrane and the endoplasmic reticulum there are pumps which transport calcium ions out of the cytosol. The result is a huge concentration difference between the cytosol on the one hand and the extracellular space and the lumen of the endoplasmic reticulum on the other hand. This can be several orders of magnitude. There are also ion channels in these membranes which, when open, allow the calcium to flow down its gradient. This provides a way to change the calcium concentration in the cytosol very fast and this can cause rapid changes in the behaviour of a cell. In this context it is important that the endoplasmic reticulum has such a high surface area and is so widely distributed in the cell. One type of calcium channels in the ER reacts to the binding of the substance IP (inositol 1,4,5-trisphosphate) to the channel by opening. This effect is also modulated by the calcium concentration in the cytosol. There are calcium channels in the plasma membrane and there is also a certain amount of leakage through both membranes. Transport of calcium in and out of mitochondria can be an important effect. Some combination of these features can lead to oscillations in the calcium concentration in the cytosol. This presents a challenge for mathematical modelling. Ideally a dynamical system consisting of ODEs for the concentrations of various substances would exhibit periodic solutions. Of course a system of this kind must have dimension at least two and several two-dimensional models have been proposed. It could be that several of these models are useful since calcium signalling in different cell types may use different mechanisms. The difficult thing is not to find a model exhibiting oscillations but to find the right model for a particular type of cell. In what follows I consider one type of model. I have chosen this type for two reasons. The first is its simplicity. The second is that it may be relevant to explaining the role of calcium in the activation of T cells.
(inositol 1,4,5-trisphosphate) to the channel by opening. This effect is also modulated by the calcium concentration in the cytosol. There are calcium channels in the plasma membrane and there is also a certain amount of leakage through both membranes. Transport of calcium in and out of mitochondria can be an important effect. Some combination of these features can lead to oscillations in the calcium concentration in the cytosol. This presents a challenge for mathematical modelling. Ideally a dynamical system consisting of ODEs for the concentrations of various substances would exhibit periodic solutions. Of course a system of this kind must have dimension at least two and several two-dimensional models have been proposed. It could be that several of these models are useful since calcium signalling in different cell types may use different mechanisms. The difficult thing is not to find a model exhibiting oscillations but to find the right model for a particular type of cell. In what follows I consider one type of model. I have chosen this type for two reasons. The first is its simplicity. The second is that it may be relevant to explaining the role of calcium in the activation of T cells.
I consider first a model due to Somogyi and Stucki (J. Biol. Chem. 266, 11068). It is a two-dimensional dynamical system. The two variables are the calcium concentrations in the lumen of the ER and the cytosol, call them  and
and 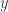 . The concentration of IP is taken to be constant. The rates of change of and are given by
. The concentration of IP is taken to be constant. The rates of change of and are given by 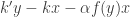 and
and  . The quantities
. The quantities  are positive constants while
are positive constants while  is a positive function which describes the behaviour of the IP receptor and must be further specified to get a definite model. The inventors of the model remark that setting
is a positive function which describes the behaviour of the IP receptor and must be further specified to get a definite model. The inventors of the model remark that setting  and
and  causes this system to reduce to the famous Brusselator, which I have commented on elsewhere. Thus the model can be thought of as a kind of generalized Brusselator and indeed it exhibits similar qualitative behaviour. The choices which are suggested to be appropriate for the cells being studied (in this case hepatocytes) is that
causes this system to reduce to the famous Brusselator, which I have commented on elsewhere. Thus the model can be thought of as a kind of generalized Brusselator and indeed it exhibits similar qualitative behaviour. The choices which are suggested to be appropriate for the cells being studied (in this case hepatocytes) is that 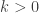 and is given by a Hill function,
and is given by a Hill function,  . Nice features of this system is that it has a unique stationary solution which can be written down explicitly and that it is also possible to get an explicit formula for the characteristic equation of the linearization at that stationary solution. In this way the stability of the stationary solution can be determined, with instability corresponding to the existence of a limit cycle. It is stated that a Hopf bifurcation occurs but there is no discussion of proving this. The general picture seems to be that oscillatory behaviour occurs at intermediate levels of IP stimulation and disappears at levels which are too low or too high. In this paper an alternative version of the model is introduced where in some places is replaced by
. Nice features of this system is that it has a unique stationary solution which can be written down explicitly and that it is also possible to get an explicit formula for the characteristic equation of the linearization at that stationary solution. In this way the stability of the stationary solution can be determined, with instability corresponding to the existence of a limit cycle. It is stated that a Hopf bifurcation occurs but there is no discussion of proving this. The general picture seems to be that oscillatory behaviour occurs at intermediate levels of IP stimulation and disappears at levels which are too low or too high. In this paper an alternative version of the model is introduced where in some places is replaced by  . This happens when modelling effects driven by the difference of concentrations in the two compartments. Given that is normally much larger than it is plausible to replace the difference of concentrations by the concentration in the ER.
. This happens when modelling effects driven by the difference of concentrations in the two compartments. Given that is normally much larger than it is plausible to replace the difference of concentrations by the concentration in the ER.
The dephosphorylation of the transcription factor NFAT during the activation of T cells has been studied in a paper of Salazar and Höfer (J. Mol. Biol. 327, 31). An important step in the activation process is an influx of calcium caused by release of IP. The calcium binds to calmodulin. It also binds to the phosphatase calcineurin which can then be activated by calmodulin. Finally calcineurin removes phosphate groups from NFAT. In this paper a model for calcium dynamics is used which is closely related to the (alternative model) of Somogyi and Stucki. There are three equations but two of them form a closed system which is more or less the Somogyi-Stucki model with a specific choice of receptor activity as a function of the concentration of IP. The last equation essentially means that the calcium level is integrated in time to give the concentration of active calcineurin.
This entry was posted on November 19, 2011 at 11:59 am and is filed under dynamical systems, immunology, mathematical biology. You can follow any responses to this entry through the RSS 2.0 feed.
You can leave a response, or trackback from your own site.

December 6, 2011 at 11:59 am |
I now noticed that in order to get the Brusselator it is necessary to take instead of the function I gave.
instead of the function I gave.
January 6, 2012 at 6:19 am |
[…] signalling pathway, its connection to calcium and a paper on the subject by Salazar and Höfer in a previous post. Now I have written a paper where I look into mathematical aspects of the activation of NFAT by […]