These days reproduction numbers for epidemiology are prominent in the popular media. Many people are familiar with the idea that stopping a resurgence of COVID-19 infections in a region has to do with making and keeping something called 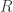 less than one. They may also be familiar with the informal definition of that it is the number of new infections caused by an infected individual. But how is (or
less than one. They may also be familiar with the informal definition of that it is the number of new infections caused by an infected individual. But how is (or 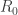 as it is more commonly called by scientists) defined? A mathematician might not expect to find an answer in the media but it might be reasonable to expect one in the scientific literature on epidemiology. In the past I have been frustrated by the extent to which this fails to be the case. What is typically given is a description in words which I never found possible to convert into a precise mathematical account, despite considerable effort. Now, in the context of a project on hepatitis C which I have been working on with colleagues from Cameroon, my attention was drawn to a paper of van den Driessche and Watmough (Math. Biosci. 180, 29) which contains some answers of the type I was looking for. I was vaguely aware of this paper before but I had never seriously tried to read it because I did not realise its nature.
as it is more commonly called by scientists) defined? A mathematician might not expect to find an answer in the media but it might be reasonable to expect one in the scientific literature on epidemiology. In the past I have been frustrated by the extent to which this fails to be the case. What is typically given is a description in words which I never found possible to convert into a precise mathematical account, despite considerable effort. Now, in the context of a project on hepatitis C which I have been working on with colleagues from Cameroon, my attention was drawn to a paper of van den Driessche and Watmough (Math. Biosci. 180, 29) which contains some answers of the type I was looking for. I was vaguely aware of this paper before but I had never seriously tried to read it because I did not realise its nature.
The context in which I would have liked to find answers is that of models given by systems of ordinary differential equations where the unknowns are the numbers of individuals in different categories (susceptible, infected, recovered etc.) as functions of time. How the numbers reported in the media are calculated (on the basis of discrete data) is something I have not yet tried to find out. At the moment I would like an answer in the context which bothered me in the past and this is the context treated in the paper mentioned above. A typical situation is that found in the basic model of virus dynamics, a system of three ODE describing the dynamics of a virus within a host, with the unknowns being uninfected cells, infected cells and virions. There is a quantity which can be expressed in terms of the coefficients of the system. If  then the only non-negative steady state is virus-free. This is the uninfected state and it is globally asymptotically stable. If
then the only non-negative steady state is virus-free. This is the uninfected state and it is globally asymptotically stable. If  there is an uninfected state which is unstable and an infected state which is positive and globally asymptotically stable. This kind of situation is not unique to this example and similar things are seen in many models of infection. There is a reproductive number (or perhaps more than one) which defines a threshold between different types of late-time behaviour.
there is an uninfected state which is unstable and an infected state which is positive and globally asymptotically stable. This kind of situation is not unique to this example and similar things are seen in many models of infection. There is a reproductive number (or perhaps more than one) which defines a threshold between different types of late-time behaviour.
It is not obvious that the analysis of van den Driessche and Watmough applies to models of in-host dynamics of a pathogen since it is necessary to make a choice of infected and uninfected compartments which is related to the biological interpretation of the variables and not just to the mathematical structure of the model. Their analysis does apply to the basic model of virus dynamics if the infected compartments are chosen to be the infected cells and virions and the reaction fluxes are partitioned in a suitable way. The simple picture of the significance of the reproductive number given above does not always hold. There is also another scenario which can occur and does so in many practical examples and involves the notion of a backward bifurcation. It goes as follows. For sufficiently small the disease-free steady state is globally asymptotically stable but as is increased this property breaks down before 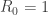 is reached. A fold bifurcation occurs which creates a stable and an unstable positive steady state. The unstable steady state moves so as to meet the disease-free state when . For there are exactly two steady states and the positive one is globally asymptotically stable. There is bifurcation for but it has a different structure from that in the classical scenario (which is a transcritical bifurcation). It bears some resemblance to a sub-critical Hopf bifurcation.
is reached. A fold bifurcation occurs which creates a stable and an unstable positive steady state. The unstable steady state moves so as to meet the disease-free state when . For there are exactly two steady states and the positive one is globally asymptotically stable. There is bifurcation for but it has a different structure from that in the classical scenario (which is a transcritical bifurcation). It bears some resemblance to a sub-critical Hopf bifurcation.
The most useful insights I got from reading and thinking about the paper of van den Driessche and Watmough are as follows. The primary significance of concerns the disease-free steady state and its stability. The fact that it can sometimes characterise the stability of an infected steady state is a kind of bonus which does not apply to all models. What it can do is to provide information about the stability of a positive steady state in a regime where it is close to the bifurcation point where it separates from the disease-free steady state. This circumstance is analysed in the paper using centre manifold theory. The significance for the stability of a steady state which is far away is a weak one. Continuity arguments can be used to propagate information about stability through parameter space but only as long as no bifurcations happen. When this is the case depends on the details of the particular example being considered. What is the definition of given in the paper? It is the largest modulus of an eigenvalue of a certain matrix (the next generation matrix) constructed from the linearization of the system about the disease-free steady state, whereby the construction of this matrix incorporates information about the biological meaning of the variables. Consider the example of the basic model of virus dynamics with the choices of infected and uninfected compartments as above. There is more than way of partitioning the reaction fluxes. I first tried to put both the production of infected cells and the production of virions into the category of fluxes called  in the paper. Applying the definition of the reproduction number given there leads to
in the paper. Applying the definition of the reproduction number given there leads to  , where is the reproduction number usually quoted for this model. If instead only the production of infected cells is put into the category then the general definition gives the conventional answer . The two quantities defining the threshold are different but the definition of being above or below the threshold are the same. (
, where is the reproduction number usually quoted for this model. If instead only the production of infected cells is put into the category then the general definition gives the conventional answer . The two quantities defining the threshold are different but the definition of being above or below the threshold are the same. ( ). That this kind of phenomenon can occur is shown by example in the paper.
). That this kind of phenomenon can occur is shown by example in the paper.
This entry was posted on June 12, 2020 at 2:56 pm and is filed under diseases, dynamical systems, mathematical biology, public health. You can follow any responses to this entry through the RSS 2.0 feed.
You can leave a response, or trackback from your own site.

November 13, 2020 at 7:57 am |
[…] was an improvement of my understanding of the concept of the basic reproductive number as discussed here and here. During this project I have learned a lot of new things, mathematical and biological, and […]
May 10, 2023 at 9:19 am |
[…] this point. There does remain a choice in splitting the positive terms, as already discussed in a previous post. It remains to consider the last condition (A5). Its interpretation in words is that if the […]