The principle of competitive exclusion arose in ecology. Roughly speaking it says that in an ecosystem with only 

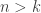
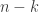
The ODE systems of relevance here are generalizations of the classical chemostat. This models a vessel containing fluid in which one species of bacteria lives. A nutrient is introduced continuously from a reservoir with fixed concentration in such a way that the rate of inflow is constant. Fluid is removed at the same rate. It is assumed that everything else the bacteria need to live and proliferate is present in sufficient quantities so as to present no limit to the population growth. The unknowns in the system are the concentrations of bacteria and nutrient. Call them 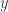
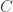




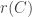
To investigate competitive exclusion it is necessary to go to models with more than one species relying on the same resource. How can periodic solutions or more complicated kinds of attractors be ruled out in that case? A recent preprint of Bernold Fiedler and Sze-Bi Hsu, available from Fiedler’s web page makes progress on understanding this question. The authors review results in which Lyapunov functions have been applied to this problem.They then introduce a new approach which is a generalization of the use of Dulac functions to systems of higher dimensions. It rules out periodic solutions for a large class of systems. What is not ruled out is orbits which, while not coming back to the same point, come back to points arbitrarily close to where they have been before. The authors appear to have ideas about how to generalize their approach to cover that case.
Competitive exclusion may also have some relevance to immunology. In a paper by De Boer and Perelson (J. Theor. Biol. 169, 375-390) they discuss the idea that T-cell clones may be selected for by competing for the chance to bind to their (suitably presented) antigen and study corresponding dynamical systems. This suggests that the T-cell response may be dependent on a sufficient variety of antigens being present.

September 8, 2011 at 6:10 pm |
[…] in a phase of exponential growth but this kind of system could also be used as a chemostat (cf. this post) to observe a steady state […]
August 25, 2012 at 9:37 am |
[…] It might be hoped that general properties of dynamical systems which arise in particular wide classes of applications might reduce some of these difficulties. An example of a class of this type is defined by the systems which describe the competition of species in ecology. In 1976 Stephen Smale (J. Math. Biol. 3, 5) showed in a three-page paper that the restriction to this type of system does not, on its own, lead to any improvement whatsoever. In more detail what he did was the following. He considered a system of ODE of the form where the are smooth and for all . The inequality expresses the competitive property while the factor on the right hand side ensures that if the population of a species is initially zero it stays zero. The region where all are non-negative, which is the biologically interesting region, is invariant. Finally he assumes that the functions are negative outside a large ball so that solutions cannot escape to infinity. Let be the set where all are non-negative and . Smale shows that there is a dynamical system in the class under discussion here where is an invariant subset, all solutions converge to as and the dynamical system on is arbitrary. Thus any complication of a general dynamical system of dimension can be built into a model of the dynamics of competing species. There is no fundamental simplification if is at least three. Soon afterwards an analogous result was proved for a class of systems in chemical physics describing continuous flow stirred tank reactors by Perelson and Wallwork (J. Chem. Phys. 66, 4390). A paper of Kaplan and Yorke (American Naturalist 111, 1030) mentions the relevance of Smale’s result to competitive exclusion, a topic mensioned in a previous post. […]